KEYWORDS
Kisspeptin, oestradiol, menstrual cycle
INTRODUCTION
Kisspeptin, discovered in 1996 in the context of tumour pathophysiology,1 is a powerful stimulator of the reproductive system. It plays a central role in influencing the timing of puberty.2 Kisspeptin is believed to be responsible for the two modes of gonadotropin-releasing hormone (GnRH) secretion: the oestrogen-induced ovulatory surge of GnRH/luteinising hormone (LH) and basal, pulsatile GnRH/LH release.3,4 Within the hypothalamus, GnRH neurons are situated very close to the kisspeptin neurons and they express kisspeptin receptor as well.5,6 Kisspeptin is believed to mediate gonadal steroid feedback to the hypothalamus. Although androgens, oestrogen and progesterone suppress gonadotropin secretion, none of these sex steroids affect GnRH secretion by direct action on GnRH neurons due to the absence of their receptors on GnRH neurons.7 Recent evidence suggests that kisspeptin neurons may act as main upstream regulators that integrate central and peripheral signals, hence causing the release of GnRH from GnRH neurons.8-10
In a female monthly cycle, just before ovulation, there is a rise in serum oestrogen which causes a rise in GnRH resulting in an LH surge. Surprisingly, GnRH neurons do not have oestrogen receptors7 whereas kisspeptin neurons do have oestrogen receptors.11 Keeping in mind the oestrogen-induced preovulatory LH surge, lack of oestrogen receptors in GnRH neurons, and their presence on kisspeptin neurons, we hypothesised that serum kisspeptin levels may vary during the various phases of the menstrual cycle and variation in these levels may have some role in ovulation. So far, virtually all research on kisspeptin signalling has focused on exogenous kisspeptin administration and its effects on reproduction. None of the studies measured fluctuations in endogenous kisspeptin secretion with a changing sex-steroid milieu. This compelled us to design the present project and to determine serum kisspeptin levels in various stages of the menstrual cycle in Saudi female students.
MATERIAL AND METHODS
Permission and ethical approval for this study were sought from the deanship of Scientific Research, University of Dammam, Saudi Arabia. Thirty female students from various colleges within our university were recruited. The study participants met the following inclusion criteria: 18-25 years old Saudi females, with a regular menstrual cycle (cycle length varying between 25 and 35 days with no more than 5 day variability), no use of prescription medications (including hormonal contraception) for at least 2 months before the study, and willing to participate. Exclusion criteria were the presence of a chronic medical condition, irregular menstruation, and inaccessibility to follow-up.
The subjects were briefed about the project in a familiarisation session. They agreed to inform us on the first day of their next cycle to plan the dates for the blood sampling. The individual length of each participant’s menstrual cycle was considered when scheduling their appointments for the experiments. Three blood samples per volunteer were collected at three different times.12
1) Early follicular phase: During days 2-5 from the onset of the menstrual cycle.
2) Preovulatory phase: During 11-16 days before the onset of the next menstrual cycle.
3) Luteal phase: 3-5 days before the onset of the next menstrual cycle.
Verification of the menstrual cycle phase was done by basal body temperature (BBT, high oestrogen during the follicular phase lowers BBT; high progesterone after ovulation raises BBT)12 and serum oestradiol levels.13-15 All blood samples were obtained by venipuncture, after an overnight fast between 8-10 a.m. (in order to minimise the effect of circadian rhythm on kisspeptin levels). These samples were allowed to clot and centrifuged within 30 minutes after venipuncture. The serum obtained was frozen at -80 °C until further analysis by ELISA kits for measurement of serum kisspeptin and oestradiol.
STATISTICAL ANALYSIS
Statistical analysis was done by SPSS version 20. Mean, maximum, minimum and standard deviation were calculated by descriptive statistics. Comparison between individual subjects during the follicular, luteal and ovulatory phase was done by one-way, repeated-measures ANOVA. In case of data violation of the assumption of sphericity, we used Greenhouse-Geisser values. To discover which specific means differed, the Bonferroni post hoc test was applied. Pearson’s correlation was used to find out the relationship. For all tests, the level of significance was set at p < 0.05.
RESULTS
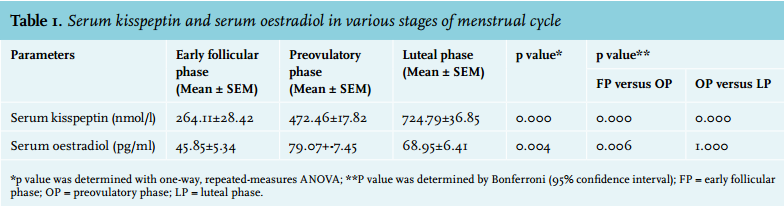
The mean duration of the menstrual cycle in all study subjects was 29±1 (mean ± SD) days (27-31 days). The duration of the menses was 6±1 days (mean ± SD). All participants recruited had, according to their past history, a regular menstrual cycle. Serum kisspeptin and oestradiol levels were different in the different phases of the menstrual cycle (table 1). Maximum concentrations of oestradiol and kisspeptin were observed in the preovulatory and luteal phase, respectively. The repeated measures ANOVA with a Greenhouse-Geisser correction revealed that mean serum kisspeptin differed statistically significantly between time points/phases (f = 61.524, p = 0.000). Post hoc testing using the Bonferroni correction revealed that there was a statistically significant increase (p = 0.000) in serum kisspeptin levels from the early follicular to the preovulatory phase (264.11±28.42 vs. 472.46±17.82 nmol/l respectively), and from the preovulatory to the luteal phase (472.46±17.82 vs. 724.79±36.85 nmol/l respectively) (table 1). Therefore, we can conclude that serum kisspeptin levels change statistically significantly across these three phases of menstruation.
The repeated measures ANOVA with a Greenhouse-Geisser correction determined that mean serum oestradiol also differed statistically significantly between time points/ phases (f = 6.632, p = 0.004). Post hoc testing using the Bonferroni correction revealed that there was a statistically significant (p = 0.006) increase in serum oestradiol levels from the early follicular to the preovulatory phase (45.85±5.34 vs. 79.07±7.45 pg/ml respectively), but statistically insignificant (p = 1.000) decrease from the preovulatory to the luteal phase (79.07±7.45 vs. 68.95±6.41 pg/ml respectively) (table 1). No correlation was found between serum kisspeptin and oestradiol in the follicular, preovulatory and luteal phase.
DISCUSSION
The present study revealed endogenous kisspeptin secretion in the various phases of the menstrual cycle in young female participants with regular menstruation. To our knowledge, this is the first analysis of its type. We did not find any association between serum kisspeptin levels and body mass index; a finding in accordance with a recent study in which no remarkable changes in kisspeptin expression were found in diet-induced obese experimental rats compared with controls.16 Serum concentrations of kisspeptin and oestradiol both increased statistically significantly in the preovulatory phase when compared with the early follicular phase (table 1). Hence, our study indicates that as the sex-steroid milieu (oestrogen and progesterone) fluctuates during the different phases of the female menstrual cycle, kisspeptin levels also fluctuate. Mammalian ovulation requires an LH surge brought about by the positive feedback action of oestradiol on GnRH release. High kisspeptin levels in the preovulatory phase suggest that kisspeptin might be responsible for the LH surge. Our results are in agreement with Smith et al., who showed that kisspeptin expression increases just before ovulation and during a steroid-induced LH surge in ovariectomised rats.17 Clarkson et al. also revealed that kisspeptin neurons become activated at the time of ovulation.18 Some studies have shown that inhibition of kisspeptin action abolishes the pro-oestrous LH surge and inhibits oestrous cyclicity in rats.19,20 Thus, kisspeptin signalling seems to be essential for the preovulatory GnRH/LH surge. As further proof in the role of kisspeptin in ovulation, recent evidence suggests that lactating rats have low levels of KiSS mRNA and protein in the hypothalamus.21 Hence, the cause of increased kisspeptin in the ovulatory phase could be increased activation of kisspeptin neurons and increased expression induced by high oestradiol as documented in above-mentioned animal studies.
Few studies have measured the response to exogenous kisspeptin administration in terms of GnRH-stimulated LH release in varying sex-steroid milieus such as the follicular, preovulatory and luteal phase women.22 Yee-Ming Chan detected that the response to kisspeptin (as measured by kisspeptin-induced, GnRH-induced LH pulse) was largest in preovulatory women, intermediate in luteal-phase women, and smallest in follicular-phase women.22 Dhillo et al. 23 similarly observed the largest LH responses in preovulatory women after exogenous administration of kisspeptin compared with the follicular and luteal phases. Jayasena et al. 24 discovered that preovulatory women had a significantly large LH response to kisspeptin but early to mid-follicular-phase women did not. These findings are also concordant with studies in rats10 and sheep25 in which the largest LH response to kisspeptin was seen just before ovulation. We also observed a significant increase in serum kisspeptin levels from the preovulatory to the luteal phase. At the moment we are unable to provide a rational clarification for this rise. However, as more and more avenues about kisspeptin are explored, we may come up with an appropriate justification in forthcoming years. The serum oestradiol levels showed typical monthly variations, with the lowest oestradiol levels in early follicular, intermediate in luteal and peak levels in preovulatory phase, in accordance with previous research.26,27 Our data also suggested that serum kisspeptin levels were not related to oestradiol in any of the stages. Further studies should be performed to explore the factors affecting serum kisspeptin levels.
CONCLUSION
By measuring serum kisspeptin levels in females during the different phases of the menstrual cycle and by relating these to oestradiol, we are beginning to gather insight into the fundamental physiology of kisspeptin across the human menstrual cycle. Our results suggest that endogenous kisspeptin secretion varies across the menstrual cycle. However, kisspeptin levels do not seem to be related to oestradiol in any of the stages. The physiological mechanisms underpinning these differences may be explored by prospective studies measuring kisspeptin under conditions where the sex-steroid milieu and other factors are directly controlled and manipulated.
STRENGT HANDLIMITATION , AND RECOMMENDATION
The strength of our study lies in its novelty, being the first one to explore kisspeptin fluctuations across the menstrual cycle. Although the sample size was relatively small, limiting our ability to draw inferences, the data are important for potential further understanding of the role of kisspeptin in the female monthly cycle. We believe that to obtain a more precise depiction of kisspeptin, the sampling regime would need to be much more frequent, and more reliable. If the role of kisspeptin in ovulation is confirmed, it might offer a potential basis for a new infertility treatment.
DISCLOSURES
The authors acknowledge the financial support from Deanship of Scientific Research, University of Dammam, Saudi Arabia by grant number 2013134.
REFERENCES